
Blog
Estadíos Larvarios de los Neurópteros Ibéricos. VI: Chrysoperla carnea (Stephens, 1836), Chrysoperla mediterranea (Hölzel, 1972) y Chrysoperla ankylopteryformis Monserrat y Díaz-Aranda, 1989. (Insecta, Neuroptera: Chrysopidae).
L. M. DIAZ- ARANDA Y V. J. MONSERRAT.
Se describen y comparan los estadios preimaginales de las tres especies ibéricas del género Chrysoperla Steinman, 1964: C. carnea (Stephens, 1836), C. mediterránea (Hólzel, 1972) y C. ankylopteryformis Monserrat y Díaz-Aranda, 1989, incluyéndose claves para la determinación de las larvas neonatas y desarrolladas. Asimismo, se aportan datos sobre la biología, etología y distribución de las tres especies citadas. L. M. Díaz-Aranda: Departamento de Biología Animal, Universidad de Alcalá. 28871 Alcalá de Henares, Madrid. V. J. Monserrat: Departamento de Biología Animal 1, Facultad de Biología, Universidad Complutense, 28040 Madrid.
Palabras clave: Chrysopidae, Chrysoperla, Estadios preimaginales, Península ibérica.
INTRODUCCIÓN
La familia Chrysopidae representa sin duda uno de los más útiles aliados del hombre en su lucha contra determinadas plagas de fitófagos, especialmente áfidos, cóccidos y aleyródidos, ya que la mayoría de las más de 1200 especies que constituyen esta familia (ASPÓCK, ASPÓCK & HÓLZEL, 1980; NEW, 1986) predan activamente sobre estos insectos, tanto en estado larvario como en
estado adulto.
Su enorme interés económico, debido a su actuación como agentes espontáneos de control, así como su empleo y cultivo por el hombre con el fin de combatir estas plagas, es sobradamente conocido y está recogido abundantemente en la bibliografía existente. A pesar de ello, la morfología de los estadíos larvarios de la mayoría de las especies descritas es aún desconocido y sólo estadios larvarios de unas 120 especies holárticas han sido estudiados y descritos con cierta profundidad (GEPP, 1984), necesitándose por ello que nuevas descripciones y datos permitan la elaboración de claves y la determinación de todas las larvas pertenecientes a las especies conocidas de crisópidos (TAUBER, 1974).
Los estadios larvarios de algunas de las 40 especies pertenecientes a la fauna ibérica han sido suficientemente descritas, pudiéndose recabar información en ASPÓCK, ASPÓCK & HÓLZEL (1980), MONSERRAT (1984a), DÍAZ-ARANDA Y MONSERRAT (1988a), MONSERRAT (1989), etc., pero aún se desconoce la morfología larvaria de muchas de ellas, por lo que resulta muy interesante aportar nuevos datos al respecto que permitan la identificación específica de estos estadíos.
En el presente artículo se aborda la descripción y comparación de los estadios preimaginales de las especies ibéricas pertenecientes al género Chrysoperla Steinman, 1964, el cual está representado en nuestra fauna por tres especies.
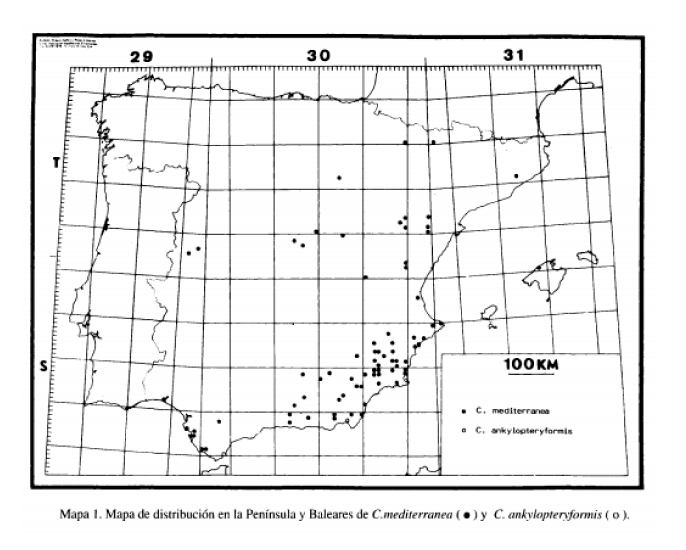
El género Chrysoperla incluye unas 25 especies y es sin duda el de mayor interés económico, habiéndose utilizado su cultivo en masa en multitud de programas de control biológico (TAUBER, 1974). Posee una distribución geográfica prácticamente cosmopolita, excepción hecha de la región australiana y es, no obstante, un género proporcionalmente escaso en número de especies, en su mayoría conocidas de la región holártica. Está representado en la región paleártica por sólo 9 especies (HÓLZEL, 1984; MONSERRAT Y DIAZ-ARANDA, 1989). Tres de ellas son conocidas en la fauna ibérica: Chrysoperla carnea (Stephens, 1836), que representa sin duda el crisópido más frecuente y de dispersión geográfica más extensa, acompañando al género al que pertenece en casi toda su amplia distribución geográfica mundial. Habita en toda la Península Ibérica y es una especie extremadamente eurioica que ocupa prácticamente todos los medios y sustratos vegetales existentes. La segunda especie ibérica es Chrysoperla mediterránea (Holzel, 1972), de distribución mediterránea occidental, mucho más termófila que la anterior, siendo por ello inexistente en las zonas peninsulares de influencia eurosiberiana y por el contrario frecuente en las zonas de mayor influencia mediterránea (Mapa n° 1). Se trata de una especie asociada a coniferas, en particular a especies del género Pinus. Por último, Chrysoperla ankylopteryformis Monserrat y Díaz-Aranda, 1989 conocida de la región árida del SE peninsular (Mapa n° 1) y de biología prácticamente desconocida.
El nivel de conocimiento sobre la morfología de los estadios preimaginales de las especies ibéricas de este género es muy variable. Así, de las dos especies más recientemente descritas, no existen datos sobre C. ankylopteryformis y son casi inexistentes los de C. mediterránea, ya que sólo DUELLI (1987) anota muy someramente datos sobre la morfología del huevo y larva de esta especie. Por el contrario, la información existente al respecto sobre C. carnea es muy abundante debido a sus características ubiquistas, a su distribución prácticamente cosmopolita y al gran interés económico que representa como aliado en la lucha contra distintos fitófagos. Muchos de estos trabajos aportan interesantes datos sobre la biología de su larva, otros aportan o ilustran datos sobre su morfología larvaria en mayor o menor profundidad, siendo de utilidad para recabar información sobre su quetotaxia, pigmentación, coloración, variabilidad, etc. (SCHNEIDER, 1851; PARISER, 1917; STEINKE, 1922; WITHYCOMBE, 1923, 1925; LACROIX, 1929; STITZ, 1931; FRANKENBERG, 1936; KILLINGTON, 1937; NEWMARK, 1952; SMIRNOFF, 1953; HOFFMANN, 1962; BANSCH; 1966; TOSCHI, 1966; AGEKJAN, 1973; TAUBER & TAUBER, 1973; EGGER, 1974; PRINCIPI & CANARD, 1974; TAUBER, 1974; ASPOCK, ASPÓCK & HOLZEL, 1980; DUELLI, 1981; GEPP, 1983; PANTALEONI, 1983; CANARD, SEMERIA «fe NEW, 1984;
etc.). Sin embargo, algunos de ellos no pasan de ser meramente testimoniales e incluso nos plantean ciertas dudas sobre su correcta asignación específica.
MATERIAL Y MÉTODO
Con el fin de estudiar todas las fases preimaginales de sus ciclos biológicos, así como de asegurar y confirmar la asignación de las larvas a describir a las tres especies anteriormente citadas, se inició la obtención de los datos a partir de hembras grávidas de las tres especies ibéricas, cuyos imagos no ofrecen ningún problema en su determinación específica (MONSERRAT Y DIAZ-ARANDA, 1989).
Las hembras de C. carnea fueron colectadas a la luz en Alcalá de Henares (Madrid) el 9. V. 1988, las de C. mediterránea fueron capturadas sobre Pinus halepensis en Valdenoches (Guadalajara) el 2. VIL 1988 y las de C. ankylopteryformis se colectaron a la luz en Rambla Morales, Cabo de Gata (Almería) el 18. VIL 1988.
Dichas hembras fueron aisladas en cajas de cultivo en cuyo interior efectuaron las correspondientes puestas, obteniéndose de ellas las larvas neonatas sobre las que iniciar el estudio morfológico.
Las larvas neonatas fueron aisladas y cultivadas en cajas de plástico transparente de 2x4x4 cm y tras la primera muda fueron transladadas a cajas cilindricas de 4.5 cm de alto x 7 cm de diámetro.
El cultivo de las larvas se desarrolló en condiciones de temperatura de 27± 1°C, con un fotoperiodo de 12:12 y una humedad relativa del 80%. Las larvas fueron alimentadas con áfidos durante todo su desarrollo, habiendo sido utilizados preferentemente Aphis fabae y A. craccivora en distintos estados de desarrollo en función del tamaño de la larva en cultivo.
Los datos de coloración y pigmentación se observaron y anotaron en vivo y para el estudio de la quetotaxia, los ejemplares fueron aclarados con ácido láctico, con el fin de mejorar la visualización de las estructuras tegumentarias.
Con el objeto de estudiar el posible grado de variabilidad de los caracteres observados, una vez estudiada y fijada la morfología de los ejemplares ex-ovo anteriormente citados, se estudiaron ejemplares de larvas en distintos estadios de C. carnea y C. mediterránea colectados por los autores en otros puntos de la Península Ibérica.
DESCRIPCIÓN DE LOS ESTADIOS PREIMAGINALES DE LAS ESPECIES IBÉRICAS DEL GÉNERO CHRYSOPERLA.
CHRYSOPERLA CARNEA (Stephens, 1836). Aspecto general según Fig. 22. A tenor de los abundantes datos existentes en la bibliografía sobre los estadios preimaginales de esta especie, la cual ha sido citada con anterioridad, nos limitaremos a aportar
los datos de su morfología (Fig. le, 5c, 10c, 14c y 16) que puedan sernos de utilidad a la hora de diferenciarla de las otras dos especies ibéricas de este género. A este respecto se harán los correspondientes comentarios en la discusión final.
CHRYSOPERLA MEDITERRANEA (Hotzel, 1972)
Huevo.
Huevo elipsoide, de color verde con micropilo blanco. Pedúnculo hialino. A medida que transcurre el desarrollo embrionario los polos adquieren tonalidad amarilla y se observan las manchas metaméricas pardas. Al final de este período los huevos son totalmente pardos, con manchas metaméricas pardo oscuras. Corion abandonado de color blanco intenso.
Ovirruptor.
La larva neonata eclosiona mediante la práctica en el corion de una fisura longitudinal que abarca desde un lateral del micropilo hasta algo más de un tercio de la longitud del huevo. El ovirruptor está esclerotizado y presenta denticulación según Fig. la.
Larva Neonata (L-l).
Longitud total del cuerpo de 2.5-3 mm. Coloración general pardo pálida, aspecto general según Fig. 2.
Cabeza de color amarillo pálido, con manchas epicraneales y genales pardo oscuras. Las primeras (Fig. 5a) abarcan desde la
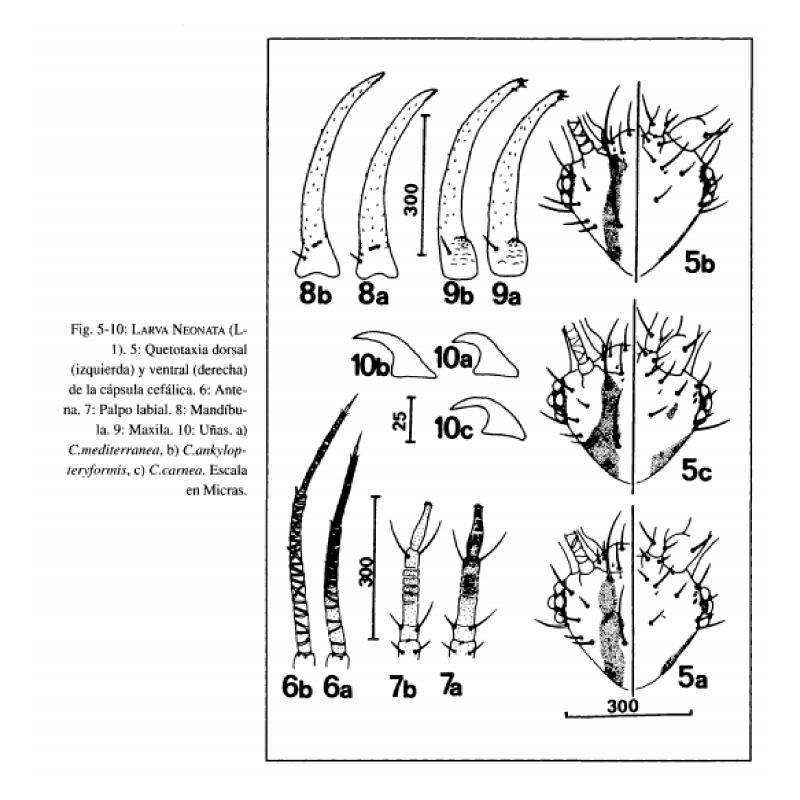
base del apodema antenal hasta el margen cervical y presentan contorno irregular y similar anchura en toda su longitud. Las segundas abarcan desde el margen cervical hasta el margen posterior de los ojos (Fig. 5a), extendiéndose ligeramente sobre la cara ventral en su margen posterior. Antenas (Fig. 6a) pardo amarillentas en su porción basal, tornándose a pardas en la porción distal. Mandíbulas (Fig. 8a) y maxilas (Fig. 9a) pardo amarillentas. Palpos labiales (Fig. 7a) pardo amarillentos en su base y pardos en la porción distal. Ojos formados por 6 estemas, 1 central y 5 periféricos, de estos últimos 3 son dorsales, 1 anteroventral y el otro posteroventral respecto al estema central. El tegumento que rodea a los estemas es de color negro. Quetotaxia cefálica dorsal y ventral según Fig. 5a
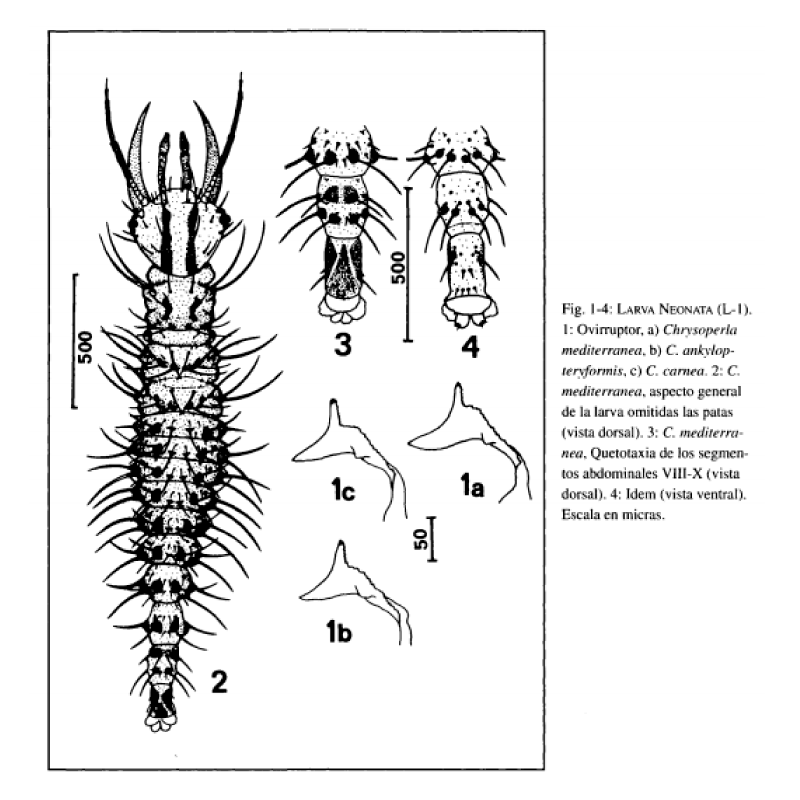
Tora (Fig. 2) de color pardo pálido, con tubérculos laterales pardos, portadores de 2 largas setas hialinas los del pronoto y 3 los del mesonoto y metanoto. Escleritos laterodorsales de los 3 segmentos torácicos pardos muy oscuros (Fig. 2). Disposición de las sedas dorsales del tórax según Fig. 2. Patas pardo pálidas, con coxa, tercio distal del fémur, tercio proximal de la tibia y tarso pardos más oscuros. Uñas según Fig. 10a.
Abdomen (Fig. 2) pardo pálido. Segmentos I-VIII con tubérculos dorsolaterales pardos, estando fuertemente esclerotizados los correspondientes a los segmentos V-VIII. Segmentos II-VIH con tubérculos laterales pardos oscuros, todos ellos fuertemente esclerotizados. Dichos tubérculos portan 1 seta hialina en los segmentos I y VIII y 2 setas hialinas en los segmentos II-VII. Quetotaxia dorsal de los segmentos abdominales I-VII según Fig. 2. Escleritos y quetotaxia dorsal y ventral de los segmentos VIIIX según Fig. 3 y 4.
Larva desarrollada (L-3).
Longitud total del cuerpo 8 – 8.5 mm. Aspecto general según Fig. 11 y 23.
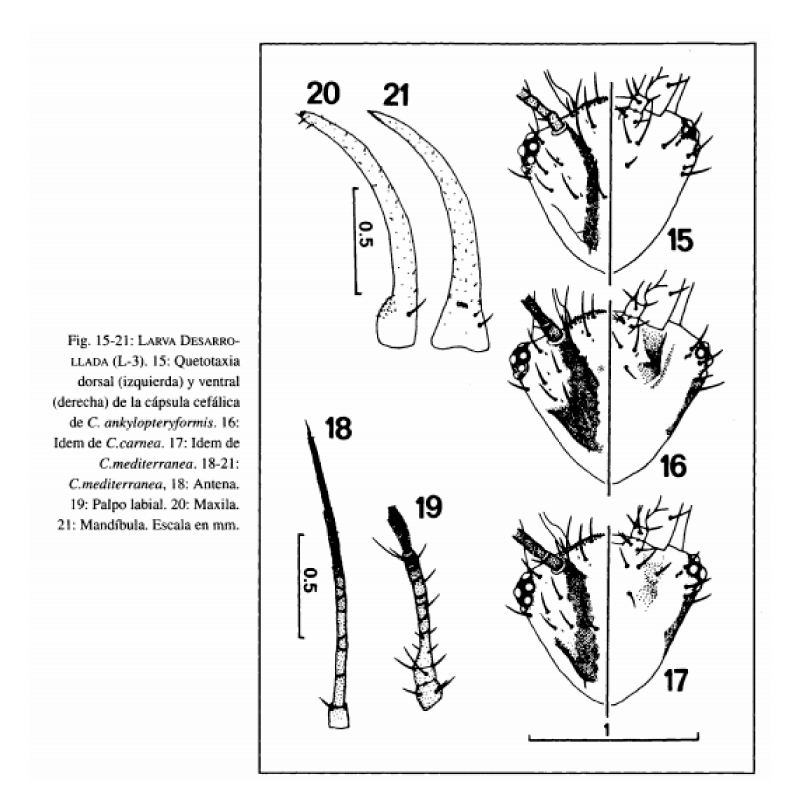
Cabeza amarillenta, con manchas epicraneales y genales pardas muy oscuras. Las primeras se extienden con similar anchura desde la base de la antena hasta ponerse en contacto con la sutura occipital, acodándose y extendiéndose levemente sobre ella (Fig. 17). Las segundas abarcan desde el margen cervical hasta el margen posterior de los ojos (Fig. 17). Antenas (Fig. 18) pardas, tornándose a pardo oscuras en la mitad distal. Mandíbulas (Fig. 21) y maxilas (Fig. 20) de color ámbar. Palpos labiales (Fig. 19) pardos, tornándose a pardo oscuro en la porción distal. Ojos formados por 6 estemas, de coloración y disposición idéntica a la indicada anteriormente para la larva neonata. Quetotaxia cefálica dorsal y ventral según Fig. 17.
Tórax amarillento, con 2 bandas dorsolaterales, una a cada lado de la línea media, de color pardo. Dichas bandas recorren los 3 segmentos torácicos en toda su longitud (Fig. 11). El mesonoto y metanoto presentan además una mancha de igual coloración bajo cada uno de los tubérculos laterales. Los tubérculos laterales del pronoto, mesonoto y metanoto son prácticamente esféricos, portadores respectivamente de 7, 9 y 9 setas hialinas. Escleritos laterodorsales de color pardo muy oscuro. Quetotaxia dorsal de los 3 segmentos torácicos (Fig. 11) con setas hialinas. Patas de color amarillento, a excepción de la coxa, tercio distal del fémur, tercio proximal de la tibia y tarso, que son de color pardo más oscuro. Uñas de color ámbar, con morfología según Fig. 14a.
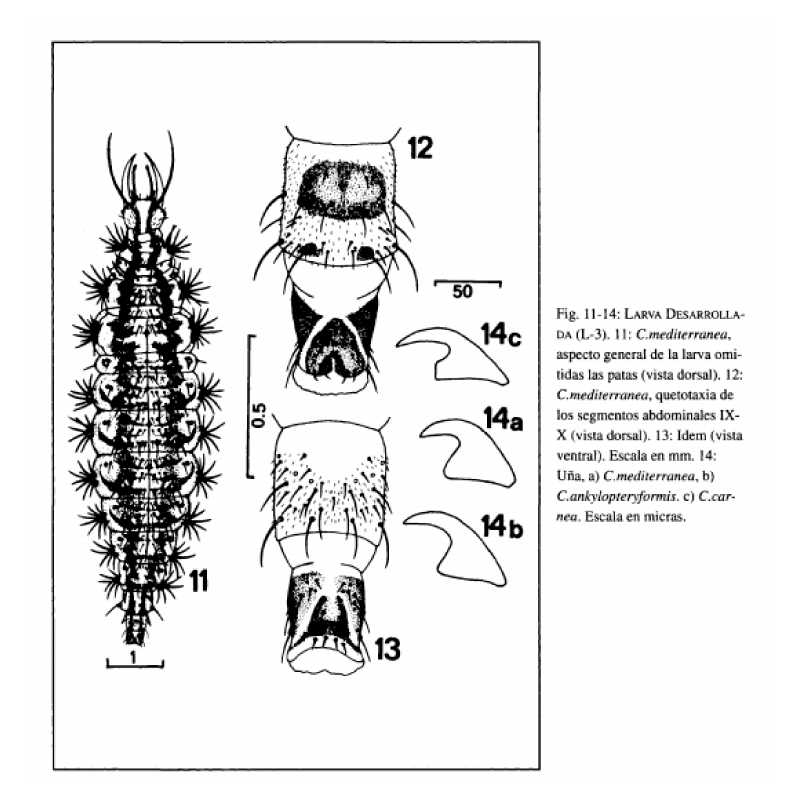
Abdomen amarillento, con 2 bandas dorsolaterales pardas, continuación de las existentes en el tórax. Dichas bandas son prácticamente continuas a lo largo de todo el abdomen, excepto en los segmentos I-V en los que no incluyen los tubérculos dorsolaterales, presentando éstos un color amarillento (Fig. 11). En cada uno de los citados segmentos, las bandas presentan un expansión lateral que parte del tubérculo dorsolateral y se dirige hasta el espiráculo (Fig. 11). En cuanto a la quetotaxia abdominal, el segmento I porta 1 pequeña seta lateral y 2 hileras de sedas dorsales. Los segmentos II-VII poseen tubérculos laterales casi esféricos, portadores de 7 a 9 setas, tubérculos laterodorsales portadores de 2 setas y 3 hileras de sedas dorsales (Fig. 11). Segmento VIII con tubérculos laterales y laterodorsales portadores de 1 seta y 2 hileras de sedas dorsales. Setas hialinas. Escleritos y quetotaxia dorsal y ventral de los segmentos IX-X según Fig. 12 y 13.
CHRYSOPERLA ANKYLOPTERYFORMIS Monserrat y Díaz-Aranda, 1989.
Huevo.
Huevo elipsoide, de color blanco verdoso, con micropilo blanco intenso. Pedúnculo hialino. A medida que transcurre el desarrollo embrionario torna a pardo, con manchas metaméricas pardo oscuras. Corion abandonado de color blanco.
Ovirruptor.
La fisura practicada en el corion por la larva neonata es prácticamente idéntica a la indicada anteriormente para C. mediterránea. Ovirruptor según Fig. Ib.
Larva Neonata (L-1).
Longitud aproximada de 3 mm. Coloración general pardo pálida. Aspecto general muy similar al de la larva neonata de C. mediterránea (Fig. 2).
Cabeza de color amarillo pálido, con manchas epicraneales y genales pardas. Las primeras abarcan desde la base del apodema
antenal hasta el margen cervical, son estrechas y presentan similar anchura en toda su longitud (Fig. 5b). Las segundas son estrechas y poco patentes, abarcando desde el margen cervical hasta el margen posterior del ojo y no se extienden apenas sobre la cara ventral (Fig. 5b). Antenas (Fig. 6b), mandíbulas (Fig. 8b), maxilas (Fig. 9b) y palpos labiales (Fig. 7b) de color pardo amarillento. Ojos idénticos a lo indicado anteriormente para C. mediterránea. Quetotaxia cefálica dorsal y ventral según Fig. 5b.
Tórax pardo pálido, los tubérculos laterales son algo más oscuros. Quetotaxia y escleritos laterodorsales prácticamente idénticos a lo indicado anteriormente para C. mediterránea (Fig. 2). Patas amarillentas, con coxa, mitad distal del fémur y tercio proximal de la tibia de color pardo. Uñas según Fig. 10b.
Abdomen pardo pálido, prácticamente igual al indicado para C. mediterránea, exceptuando que únicamente los tubérculos laterodorsales y laterales de los segmentos V-VIII están fuertemente esclerotizados, estando los de los segmentos II-IV muy ligeramente esclerotizados.
Larva Desarrollada (L-3).
Longitud aproximada de 8 – 8.5 mm. Aspecto general (Fig. 24) muy similar al de la larva de C. mediterránea (Fig. 23).
Cabeza amarillenta, con manchas epicraneales pardo oscuras, extendiéndose desde la base de la antena hasta la sutura occipital. Dichas manchas son arqueadas y estrechas, levemente ensanchadas desde su margen anterior a su extremo posterior, pero en ningún caso se extienden sobre la sutura occipital (Fig. 15). Manchas genales prácticamente inexistentes. Antenas de color pardo amarillento, ligeramente pardas en su porción distal. Mandíbulas y maxilas de color ámbar. Palpos labiales pardo amarillentos, algo más oscuros en su porción distal. Estemas de coloración y disposición idéntica a lo indicado para C. mediterránea. Quetotaxia cefálica dorsal y ventral según Fig. 15.
Tórax de color amarillo pálido, recorrido en toda su longitud por 2 bandas dorsolaterales iguales a las mencionadas para C.
mediterránea, pero más estrechas y de color rojizo. Pronoto muy ancho (Fig. 24), tubérculos laterales del pronoto, mesonoto y metanoto prácticamente esféricos, portadores respectivamente de 7, 11 y 11 setas hialinas. Disposición de las sedas dorsales de los 3 segmentos torácicos igual a la indicada anteriormente para C. mediterránea (Fig. 11). Patas amarillentas, con coxa y tarsos pardos. Uñas de color ámbar, con morfología según Fig. 14b.
Abdomen de color amarillo pálido, con 2 bandas dorsolaterales rojizas, 1 a cada lado de la línea media. Dichas bandas se hacen
difusas de los segmentos I al V, quedando así reducidas a 4 manchas en cada uno de estos segmentos, 2 a cada lado de la línea media, de las cuales una es anterior, pequeña y circular y la otra es posterior, más grande y rodea el tubérculo laterodorsal, presentando una expansión que se prolonga lateralmente hasta el espiráculo (Fig. 24). De los segmentos VI al IX las bandas vuelven ser continuas. En cuanto a la quetotaxia abdominal, el segmento I presenta 2 pequeñas setas laterales. Los segmentos II-VII poseen tubérculos laterales portadores de 8 a 10 largas setas. La disposición de las sedas dorsales de los segmentos I-VIII es igual a la indicada para C. mediterránea (Fig. 11). Setas hialinas. Escleritos y quetotaxia dorsal y ventral de los segmentos IXX casi idéntica a la ilustrada para C. mediterránea (Fig. 12, 13).
DISCUSIÓN
Al margen de los datos ya anotados anteriormente sobre las especies citadas, la información existente sobre las fases preimaginales de las especies pertenecientes al género Chrysoperla es en general poco conocida y muy dispersa. Existen datos fidedignos y otros más bien testimoniales de C. rufilabris (BURMEISTER, 1839), (SMITH, 1922, TAUBER, 1974); C. plorabunda (FITCH, 1856) (SMITH, 1922; PUTMAN, 1937; MUMA, 1959); C. externa (HAGEN, 1861) (TAUBEr, 1974); C. zastrowi (Esben-Petersen, 1928)
(Barnes, 7975); C. californica (Coq, 1890), (ESSIG, 1911); C. lanata (BANKS, 1910), (BRUCH, 1917a; Ru et al., 1975; CROUZEL & Borro, 1976; CROUZEL & SAINI, 1979); C. nipponensis (OKAMOTO, 1914), (TSUKAGUCHI, 1977); C. púdica (NAVAS, 1914), (TJEDER, 1966). etc.
No obstante, las características morfológicas correspondientes a las distintas fases larvarias del género Chrysoperla fueron bien definidas por TAUBER (1974) para especies norteamericanas, estas características pueden hacerse perfectamente extensibles a las especies ibéricas aquí descritas. Al margen de la escasa variabilidad instraespecífica observada por los autores en el material estudiado y de la anotada en la bibliografía citada, en particular en TAUBER & TAUBER (1973) y TAUBER (1974), puede considerarse al género Chrysoperla como un género bien definido desde el punto de vista de la morfología, setación y quetotaxia larvaria. Aparentemente todas las especies conocidas del género siguen un patrón bien delimitado y, en general, diferente al existente en otros géneros de la familia Chrysopidae.
Circunscribiéndonos a las especies ibéricas de este género y debido a la citada homogeneidad morfológica, las fases larvarias de C. carnea, C. mediterránea y C. ankylopteryformis son muy similares entre sí (Fig. 22, 23 y 24). Sin embargo, un examen más detallado de algunos de los caracteres indicados para las larvas completamente desarrolladas de estas tres especies, nos permiten diferenciarlas inequívocamente.
En relación a las manchas cefálicas epicraneales, cuyo interés taxonómico ya ha sido anotado por TAUBER & TAUBER (1973) y TAUBER (1974), las de C. ankylopteryformis son muy estrechas y nunca se extienden a lo largo de la sutura occipital (Fig. 15), en C mediterránea son más anchas, de contorno más irregular y se acodan levemente sobre la citada sutura (Fig. 17). Por último, en C. carnea estas manchas adquieren un aspecto más triangular al extenderse y continuarse a lo largo de toda la sutura occipital (Fig. 16), esta característica ya había sido anotada por PARISER (1917), WITHYCOMBE (1923), KILLINGTON (1937), TAUBER & TAUBER (1973), TAUBER (1974), GEPP (1983), etc.
Por otro lado, sólo C. carnea posee mancha frontoclipeal, en distinto grado de desarrollo. Así, entre el material estudiado se han observado ejemplares que poseen desde una simple sombra a uno y otro lado de la línea media (Fig. 16) a una patente mancha oscura a ambos lados de la línea media, que pueden en algunos ejemplares estar fusionadas sobre ella, de forma similar
a lo indicado para esta especie por SCHNEIDER (1851), PARISER (1917), WITHYCOMBE (1923), KILLINGTON (1937), SMIRNOFF (1953), PRINCIPI & CANARD (1974), GEPP (1983), etc. En cualquier caso, esta mancha cefálica nunca ha sido observada en C. mediterránea y C. ankylopteryformis.
Las manchas genales y ventrales de la cápsula cefálica siempre están presentes en C. carnea y C. mediterránea en distinto grado de desarrollo (Fig. 16, 17), pero en C. ankylopteryformis las genales son muy poco patentes y las ventrales son inexistentes (Fig. 15).
Todas las características de pigmentación cefálica anteriormente anotadas son prácticamente extensibles a lo observado en las larvas neonatas de estas especies (Fig. 5a, 5b, 5c).
El segundo carácter digno a tener en cuenta es la morfología de las uñas, que hasta ahora no había sido empleado en la distinción específica de las larvas de este género y que, en el caso que nos ocupa, este carácter se mantiene en los imagos de las especies citadas. Así, las larvas del tercer estadio de C. carnea poseen uñas dilatadas y acodadas en su base, dándole un aspecto cuadrangular (Fig. 14c). Por el contrario, en C. ankylopteryformis y C. mediterránea las uñas, a pesar de estar dilatadas en su base, carecen de este proceso, dándoles un aspecto triangular (Fig. 14a, 14b). En C. mediterránea el margen interno de la parte basal de la uña se acoda marcadamente con el margen interno de la parte distal (Fig. 14a), mientras que en C. ankylopteryformis ambos márgenes se disponen de forma más continuada.
Estos caracteres, observados en las larvas desarrolladas, se encuentran intuidos en las fases neonatas (Fig. 10a, 10b, 10c).
Por último, deben anotarse ciertos datos en relación a la quetotaxia torácica y abdominal. Así, BRUCH (1917), TAUBER (1974) y Ru et al., (1975) observaron que el número de setas de los tubérculos laterales de pronoto, mesonoto y metan oto es de 7, 9 y 9 respectivamente y el número de setas de los tubérculos laterales de los segmentos abdominales II-VII es de 7 a 9 para las especies americanas. Estos datos se cumplen en los ejemplares de C. carnea y C. mediterránea estudiados, no así en C. ankylopteryformis, cuyo número de setas es de 7, 11 y 11 para los tubérculos laterales del pronoto, mesonoto y metanoto respectivamente y de 8 a 10 setas para los abdominales. Así mismo, las setas en C. mediterránea y C. ankylopteryformis son hialinas, mientras que en C. carnea son ahumadas.
Otros caracteres anteriormente comentados, de morfología y pigmentación de menor importancia (como la anchura del
pronoto o el color de las bandas dorsolaterales del tórax y abdomen) han sido estudiados en las tres especies, que junto a
los 3 caracteres más relevantes de pigmentación cefálica, morfología de las uñas y quetotaxia torácica y abdominal anteriormente mencionados, permiten identificar a las larvas de las tres especies ibéricas de este género.


CLAVE PARA LA DETERMINACIÓN DE LAS LARVAS NEONATAS (L-l) DE LAS ESPECIES IBÉRICAS DEL GÉNERO CHRYSOPERLA.
1) a) Mancha epicraneal de aspecto triangular, ensanchada en su margen posterior sobre la sutura occipital (Fig. 5c). Porción
distal de las uñas muy curvada sobre su porción basal (Fig. 10c) C. carnea
b) Mancha epicraneal no triangular, ni ensanchada posteriormente sobre la sutura occipital (Fig. 5a, 5b). Porción distal de las
uñas levemente curvada sobre su porción basal (Fig. 10a, 10b) 2.
2) a) Mancha epicraneal estrecha y arqueada (Fig. 5b). Antenas y palpos labiales hialinos (Fig. 6b, 7b). Uñas con porción distal muy desarrollada (Fig. 10b) C. ankylopteryformis.
b) Mancha epicraneal ancha y de contorno irregular (Fig. 5a). Antenas y palpos labiales pardos en su porción distal (Fig. 6a, 7a). Uñas con la porción distal poco desarrollada (Fig. 10a) C. mediterránea.
CLAVE PARA LA DETERMINACIÓN DE LAS LARVAS DESARROLLADAS (L-3) DE LAS ESPECIES IBÉRICAS DEL GÉNERO CHRYSOPERLA
1) a) Uñas con porción basal acodada en su margen interno, dándole un aspecto cuadrangular (Fig. 14c) C. carnea.
b) Uñas con porción basal no acodada en su margen interno, dándole un aspecto triangular (Fig. 14a, 14b) 2.
2) a) Mancha epicraneal estrecha y no extendida sobre la sutura occipital (Fig. 15), tubérculos laterales del pronoto, mesonoto y
metanoto con 7, 11 y 11 setas respectivamente. Tubérculos laterales de los segmentos abdominales II-VII portadores de 8 a 10
setas. Uñas con la porción basal del margen interno no acodada con la porción distal (Fig. 14b) C. ankylopteryformis.
b) Mancha epicraneal ancha, acodada y extendida sobre la sutura occipital (Fig. 17).
Tubérculos laterales del pronoto, mesonoto y metanoto con 7, 9 y 9 setas respectivamente. Tubérculos laterales de los segmentos abdominales II-VII portadores de 7 a 9 setas. Uñas con la porción basal del margen interno acodada con la porción distal (Fig. 14a) C. mediterránea.
BIOLOGÍA Y COMPORTAMIENTO
Las puestas fueron realizadas de manera idéntica en las tres especies estudiadas. Los huevos fueron depositados individualmente (no agrupados), indistintamente en la pared y techo de las cajas de cultivo. El comportamiento de las larvas es también muy similar. Son larvas desnudas, que no se cubren con restos de presas y durante todo su desarrollo, se caracterizan por su rápidos movimientos y enorme voracidad, habiéndose observado canibalismo en los ejemplares de las tres especies sometidos a falta de alimento. El comportamiento en la captura de presas es básicamente el mismo al indicado para C. carnea en los trabajos de ALROUECHDI (1981), CANARD, SEMERIA & NEW (1984), etc.
Los datos obtenidos sobre la duración media de cada uno de los estadios preimaginales pueden recabarse en el cuadro 1. Los datos obtenidos en C. carnea son muy similares a los anotados por otros autores para esta especie (TOSCHI, 1966; BUTLER & RITCHIE, 1970; PASQUALINI, 1975; ALROUECHDI & VOEGELE, 1981; etc), si bien, los estadios L-2 y L-3, parecen presentar una duración más corta en los ejemplares estudiados por nosotros que en los estudiados por los autores anteriormente citados. Respecto a la duración de los estadios preimaginales de C. mediterránea y C. ankylopteryformis, no hay datos al respecto en la bibliografía existente, pero los valores obtenidos por nosotros se mantienen dentro del rango anotado para otras especies del género Chrysoperla (BURKE & MARTIN, 1956; TAUBER & TAUBER, 1974; Ru et al., 1975; BARNES, 1975; CROUZEL & Borro, 1976; BRETTELL, 1982, etc).
En cuanto a la biología del imago de las tres especies estudiadas, C. carnea es extremadamente eurioica, ocupando prácticamente todos los medios vegetales existentes en la Península Ibérica. Sin embargo, C. mediterránea, de tendencias más termófilas, parece estar fuertemente asociada a coniferas del género Pinus, especialmente a P. halepensis, donde se han contabilizado la mayoría de las capturas, tanto en estado de imago como de larva (HÓLZEL & OHM, 1972; MONSERRAT, 1985, 1987; DIAZ-ARANDA Y MONSERRAT, 1988b; DUELLI, 1987; etc.). Otras coniferas donde ha sido colectada, aunque en menor medida, son P. pinaster, P. nigra, Juníperas oxycedrus, J. thurifera y Cedras serpenvirens. Además, existen capturas puntuales sobre Tamarix gallica, Ceratonia siliqua, Quercus rotundifolia, Castanea sativa, Ilex aquifolium y en trampa de luz incandescente.
Por último, sobre la biología de C. ankylopteryformis carecemos prácticamente de datos, pareciendo estar asociada a las ramblas pedregosas de las zonas áridas del SE peninsular (MONSERRAT y DIAZ-ARANDA, 1989).

DISTRIBUCIÓN GEOGRÁFICA
Como se indicó anteriormente, C. carnea ínsula Ibérica y Baleares, abarcándola en su totalidad y existiendo multitud de citas sobre
su distribución geográfica en nuestra fauna. Es por esta razón por la que nos abstenemos de anotar tales referencias y en el mapa de distribución sólo marcaremos los puntos geográficos correspondientes a las capturas y citas de las otras dos especies ibéricas.
De C. mediterránea, con tendencias más termófilas, sólo existen citas en la Península Ibérica en zonas de influencia mediterránea (HÓLZEL, 1972; HÓLZEL & OHM, 1972; ASPÓCK, ASPÓCK & HÓLZEL, 1980; MONSERRAT, 1978, 1980, 1982, 1984b, 1984c, 1985, 1987; MONSERRAT y DIAZ-ARANDA, 1987, 1988, 1989; DIAZ-ARANDA y MONSERRAT, 1988b, 1988c, 1988d; DIAZ-ARANDA, MONSERRAT y MARÍN, 1986) (Mapa 1).
Por último, C. ankylopteryformis (Mapa 1), solamente ha sido citada hasta ahora en el SE peninsular (MONSERRAT Y DIAZARANDA, 1989).
AGRADECIMIENTO
Para la realización de este trabajo se ha contado con fondos del proyecto PB85/0236 de la C. I. C. Y. T.
