
Blog
Enzimas y toxinas de hongos entomopatógenos, su aplicación potencial como insecticidas y fungicidas
Karina Guadalupe Franco Chávez, Silvia Rodríguez Navarro, José Francisco Cervantes Mayagoitia, Juan Esteban Barranco Florido
Resumen. Los hongos entomopatógenos (HE) al infectar a los insectos producen metabolitos, como las enzimas hidrolíticas y los depsipéptidos cíclicos. Estas enzimas degradan la cutícula del insecto, pero también pueden afectar la pared celular de los hongos, similar al antagonismo de Trichoderma harzianum y de otros microorganismos utilizados como fungicidas. Los depsipéptidos tienen propiedades insecticidas y de actividad antialimentaria que ocasionan la muerte de insectos. Además, estos hongos al encontrarse en nichos ecológicos con alta biodiversidad y competencia sintetizan antibióticos y agentes fungicidas que afectan la síntesis de la pared celular de los hongos, estos metabolitos son equinocandinas, polioxinas y nicomicinas. Todos los compuestos bioactivos señalados pueden ser utilizados como bioinsecticidas o fungicidas, y ser producidos por procesos fermentativos. Esto abre nuevas áreas de desarrollo agrobiotecnológico, acorde con las nuevas tendencias de la investigación agrícola a nivel mundial.
Palabras clave: Hongos entomopatógenos, enzimas hidrolíticas, depsipéptidos, antibióticos.
INTRODUCCIÓN
Los microorganismos han sido utilizados para el control biológico de insectos y enfermedades de plantas desde principios del siglo pasado, y en las dos últimas décadas se han demostrado las ventajas del uso de los microorganismos frente al manejo de insecticidas químicos, ya que éstos generan graves problemas como daños al ecosistema, afectación a la salud humana y la aparición de resistencia en muchas especies de insectos. La solución ha sido incrementar la dosis y disminuir tiempos de aplicación del insecticida químico con el daño consecuente al medio ambiente. La presencia de insectos resistentes a los químicos y la aparición de enfermedades en las plantas, ocasionadas por los hongos, ha producido enormes pérdidas agrícolas en México, en el caso del estado de Guanajuato se pierde 22% del maíz almacenado (García-Leaños et al., 2007). Sus efectos inmediatos son el incremento de la dependencia alimentaria del país, así como su impacto en la exportación de productos agrícolas por las restricciones dadas en la Unión Europea y en Estados Unidos respecto a los químicos residuales en vegetales y frutas (Hussey y Bell, 2004).
Los principales grupos de microorganismos utilizados para el control biológico son los virus, bacterias, hongos y protozoarios. En el caso de los hongos entomopatógenos (HE), éstos han sido estudiados y aplicados en el mundo por su eficiencia en matar a los insectos, mantenerse un tiempo largo en el campo después de su aplicación, por su interacción específica con el insecto plaga y por ser relativamente seguros respecto al medio ambiente. En México, el uso de HE no se ha generalizado, aunque el empleo de dichos microorganismos para el combate de plagas se inició desde hace 50 años, sin embargo, se ha venido incrementando de forma significativa a partir de 1990 (Tamez-Guerra et al., 2001). El mecanismo de patogenicidad de los hongos entomopatógenos ha sido intensamente estudiado y se conocen las etapas principales del proceso infectivo, lo que ha permitido poder establecer estrategias para mejorar su aplicación como control biológico (Srivastava et al., 2009).
En general, el mecanismo inicia con la adhesión de los conidios sobre la superficie del exoesqueleto, proceso que sugiere una interacción entre las hidrofobinas que se encuentra en el conidio y la superficie hidrofóbica del exoesqueleto del insecto susceptible (Fang et al., 2007). Posteriormente, germina y dependiendo de la especie de hongo forma el apresorio o una sustancia mucilaginosa que lo mantiene unido para iniciar la formación del túbulo germinal que penetra por los poros o las capas externas de la epicutícula (Shah y Pell, 2003). Recientemente se ha relacionado la degradación de los lípidos de la capa externa como una etapa de crecimiento previo a la penetración del hongo (Beys da Silva et al., 2010). Se ha sugerido la posterior síntesis de enzimas hidrolíticas (proteasas, quitinasas y lipasas, entre otras reportadas) que degradan la cutícula y liberan nutrientes para el hongo, en esta etapa presenta un crecimiento micelial. Al llegar a la hemolinfa se transforman morfológicamente en células individuales denominadas blastosporas, en esta etapa se activan los mecanismos de defensa del insecto, y el hongo debe superarlos para completar el proceso de infección con la síntesis de otro tipo de proteasas que degradan el sistema inmune humoral, también de depsipéptidos cíclicos como las destruxinas que causan parálisis al insecto por su habilidad de abrir los canales de calcio (Pal et al., 2007), así como otras toxinas que dañan el sistema muscular y los tubos de Malpighi, afectando la excreción y dificultando su capacidad de alimentarse y moverse (Pal et al., 2007), para finalmente ocasionar la muerte del insecto. Posteriormente, invade totalmente al insecto y emerge en forma de hifa, esporula y sintetiza antibióticos para evitar el crecimiento de microorganismos oportunistas e inicia otro ciclo de infección (Lee et al., 2005; Srivastava et al., 2009). Con los HE se han desarrollado una gran cantidad de productos que se venden comercialmente a nivel mundial, mientras que en México está limitada a pocos productos como BEA-SIN®, PAE-SIN®, META-SIN® y VERTA-SIN® de Agrobionsa (Torres et al., 2007). Las formulaciones tienen como principio activo esporas (conidios) y lo único en que varían es el vehículo inerte que protege a la espora de los efectos de los rayos ultravioleta y mantiene la humedadrequerida. Un espacio de oportunidad en investigación poco desarrollado aún, corresponde a las enzimas y metabolitos producidos por los HE con una aplicación dual como bioinsecticidas y fungicidas, en esta revisión se documentan algunas propiedades que tienen estos metabolitos y que abren la posibilidad de una nueva área de investigación para este grupo de hongos.
La revisión se enfoca inicialmente en las enzimas (proteasas, quitinasas y glucanasas) que sintetizan los HE y que degradan la cutícula del insecto y la pared celular de los hongos, como productos con actividad biológica contra insectos y hongos. Posteriormente se referirá a los metabolitos que producen los HE y que muestran actividad insecticida y fungicida.
ENZIMAS
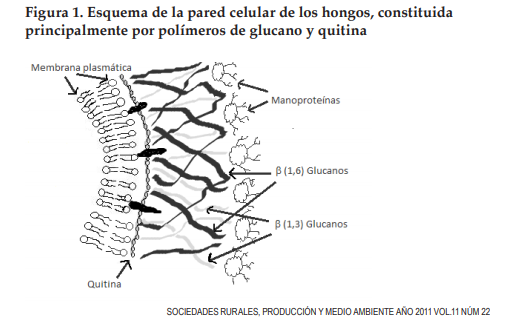
La cutícula de los insectos es una estructura consistente en nanofibras de quitina cristalina integrada a una matriz de proteínas, polifenoles, agua y pequeñas cantidades de lípidos (Vincent y Wegst, 2004), la cual puede ser degradada por proteasas y por quitinasas. Mientras que la pared celular de los hongos es una estructura compleja compuesta de quitina, glucanos y otros polímeros (Figura 1), con evidencias de que se forma un entrecruzamiento entre estos componentes, creando una compleja estructura reticular. Identificándose a las quitinasas y glucanasas como las hidrolasas que degradan la pared celular de los hongos (Adams, 2004). Por lo que las enzimas de HE que hidrolizan estas estructuras pueden ser empleadas con fines de control biológico.
Las proteasas se clasifican en cuatro diferentes clases: serina-, cisteína-, metalo- y aspartil-, de las cuales la más común en los HE son las serina-proteasas del tipo subtilisina (St Leger, 1995). También producen proteasas del tipo tripsina denominadas PR2 (Cole et al., 1993).
Ambas proteasas son enzimas extracelulares (Tiago et al., 2002) secretadas eficientemente para la degradación de la cutícula, proporcionando nutrientes para el crecimiento del hongo. La síntesis de proteasas forma parte de los procesos infectivos en los hongos patógenos Aspergillus fumigatus, Candida albicans, Coccidioides immitis y Aspergillus niger (Monod et al., 2002); es posible utilizar de la misma manera las proteasas producidas por los hongos entomopatógenos en el control de insectos, hongos fitopatógenos, bacterias y nematodos (Khan et al., 2003), las estrategias que se han sugerido, entre otras, han sido la modificación genética de Metarhizium anisopliae para sobreexpresar la proteasa PR1 (St. Leger et al., 1996), principal enzima que degrada la cutícula. Al diseminarse en el campo se demostró que no afectaba a otros insectos como el escarabajo Calosoma sycopantha, depredador de orugas y los estafilínidos, manteniendo su estabilidad genética durante un año y sin afectar antagónicamente con el complejo y dinámico microambiente de la rizósfera (Hu y St. Leger, 2002); otra estrategia es hacer crecer a los hongos utilizando inductores que permiten una eficiente producción de estas enzimas en cultivos líquidos o sólidos (Pereira et al., 2007) y que permiten mantener la virulencia de los HE. Sin embargo, las quitinasas que actúan sinérgicamente con las proteasas y las β-1,3 glucanasas son las enzimas que están involucradas directamente en la degradación de la cutícula de los insectos plaga (St-Leger et al., 1986), además de estar implicadas en muchas etapas como la germinación, el crecimiento hifal, la morfogénesis, la nutrición y la defensa contra competidores (Seidl, 2008). Respecto a lo último se ha demostrado que estas enzimas degradan la pared celular de los hongos fitopatógenos (Gohel et al., 2006). Muchas especies de bacterias, hongos y plantas producen enzimas quitinolíticas que están clasificadas en tres tipos: exoquitinasas, endoquitinasas y quitobiosas, las cuales varían entre cada especie, dependiendo del número, tipo y posición de los dominios catalíticos y de unión al sustrato (Kobayashi et al., 2002).
Otro ejemplo de la acción antifúngica de las quitinasas son las enzimas producidas por la levadura Tilleptiosis palescens y por el hongo Gliocadium catenulatum por su actividad patogénica sobre Podosphera xanthii y Pythium aphanidermatum, respectivamente, hongos causantes de enfermedades en las plantas (Punja y Utkhede, 2003). Someya et al. (2001) propusieron combinar sinérgicamente la actividad antifungicida de las enzimas quitinolíticas con el metabolito secundario prodigiosina, ambos producidos por Serratia marcescens para controlar al hongo fitopatógeno Botrytis cinerea. Por lo que se sugiere utilizar los HE como agentes fungicidas dada su capacidad de sintetizar enzimas quitinolíticas (Shubakov y Kucheryavykh, 2004). Respecto al control de insectos, se incrementó la virulencia de los HE sobre áfidos (Myzus persicae), transformando una cepa de B. bassiana con la construcción de un gen fusionado que contenía el gen Bbchit1 y el gen Pr1A que codifican a una quitinasa de 33 kDa y a una proteasa del tipo subtilisina, respectivamente; al expresarse dicho gen, la proteína se disociaba en dos partes que presentaron actividad de quitinasa y proteasa (Fang et al., 2009).
Las glucanasas degradan los β-1,3 glucanos, principales componentes de la pared celular de los hongos, ya que se producen aberturas por las cuales los hongos obtienen nutrientes; se han reportado múltiples glucanasas en hongos tales como Acremonium sp (Jayus et al., 2004) y en Trichoderma harzianum (Almeida et al., 2007), este último ha sido estudiado ampliamente por su papel como antagonista de hongos fitopatógenos (Howell, 2003). Se ha observado que las glucanasas pueden ser inducidas por la presencia de β-1,3 glucanos de paredes celulares de Rhizoctonia solani (Bara et al., 2003), sin embargo, sustratos diferentes como quitina, quitosano y N-acetilglucosamina pueden ser inductores de glucanasas (Noronha et al., 2000). Por tanto las proteasas, quitinasas y glucanasas pueden ser inducidas por sustratos complejos como la cutícula de los insectos o la pared celular de los hongos o a partir de sustratos simples, algunos estudios han demostrado este hecho, como la Exo-α-1,3-glucanasa (AGN13.1), inducida por quitina en T. harzianum (Ait-Lahsen et al., 2001); la serina proteasa P32 del hongo nematófago Verticillium suchlasporium, inducida por la cutícula de Callosubruchus maculatus (Tikhonov et al., 2002), y las proteasas y quitinasas que sin tetiza Metarhizium anisopliae en presencia de cutícula de Callosobruchus maculatus (Murad et al., 2006), esa capacidad que tienen los hongos para inducir la expresión de las enzimas con diferentes sustratos o moléculas abre la posibilidad para utilizarse tanto en el control de plagas de insecto, como en el control de enfermedades en plantas ocasionadas por hongos.
El hecho de que los HE tengan la capacidad de sintetizar las enzimas que degradan la pared celular de los hongos los hacen candidatos para tener una función antifúngica y tal vez micoparasitaria, como lo demuestran los hongos endofíticos Choiromyces aboriginum, Stachybotrys elegans y Cylindrocarpon sp (Cao et al., 2009), la bacteria Bacillus licheniformis cepa MY75 (Xiao et al., 2009) y la actinobacteria Streptomyces sp (Quecine et al., 2008). Sin embargo, es escasa la literatura en la que se propongan el uso de HE, sólo se ha mencionado a Verticillium lecanii en el control del hongo Sphaerotheca fuliginea (Askary et al., 1997).
METABOLITOS
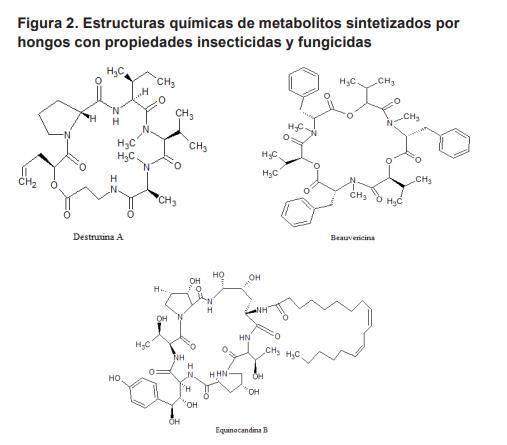
Los HE sintetizan metabolitos que son tóxicos contra insectos y generalmente son sintetizados cuando el hongo ha penetrado el exoesqueleto (Tellez-Jurado et al., 2009) y ha alcanzado el hemocele como parte de los factores de virulencia durante la patogénesis, por lo que se consideran que tienen propiedades insecticidas (Vey et al., 2001), así como de otros metabolitos con actividad antialimentaria en los insectos (Quesada-Moraga et al., 2006). Muchos de esas toxinas fúngicas son metabolitos secundarios de bajos pesos moleculares con un potencial comercial como agentes de control de insectos. Los depsipéptidos son los metabolitos mas conocidos, la denominación depende del hongo al que corresponda, por ejemplo: destruxina si es producida por Metarhizium anisopliae o beauvericina si es de Beauveria bassiana (Figura 2). Químicamente los depsipéptidos cíclicos poseen un α hidroxiácido y cinco residuos de aminoácidos, en el caso de las destruxinas se han aislado cinco análogos, denominados A a la E (Krasnoof et al., 1996; Liu et al., 2000) y que difieren en el grupo –R de los residuos hidroxiácidos, y se han identificado 35 análogos de otras especies de hongos entomopatógenos. Estas toxinas tienen un amplio rango de efectos biológicos en los insectos, entre los que se incluyen: el inducir la despolarización de la membrana debido a la apertura de los canales de Ca2+, causando la parálisis tetánica y muerte (Samuels et al., 1988), también causan cambios morfológicos y del citoesqueleto de los plasmocitos del insecto in vitro, que afecta parte de la respuesta inmune como la encapsulación y la fagocitosis (Veyet al., 2002). Además de reducir la expresión de péptidos antimicrobianos que tienen un papel importante en la respuesta inmune humoral de los insectos (Pal et al., 2007), inducen también cambios estructurales en las células epiteliales que ocasionan la disrupción de la membrana,
así como un estrés oxidativo en las células (Sowjanya y Padmaja, 2008); también inhiben la tasa de secreción de fluidos en los tubos de Malpighi (Ruiz-Sanchez et al., 2010). El efecto tóxico de los depsipéptidos en diferentes especies de insectos ha abierto una gran oportunidad para su producción como insecticida por fermentación líquida (Liu et al., 2000), encontrándose que su producción depende fuertemente de la disponibilidad de nitrógeno (Wang et al., 2009). Uno de los problemas en estos sistemas es la formación de pellets, que son núcleos densos de hifas empacadas donde el grosor de la capa externa de crecimiento sería la limitante principal para la tasa de difusión de los nutrientes (Verma et al., 2001) y donde el tamaño del pellet formado afecta la producción de
los depsipéptidos (Feng et al., 2004).
Otros metabolitos como las beauverolidos producidos por Beauveria tenella y Paecilomyces fumosoroseus no tienen efecto insecticida de manera directa, sin embargo, estos metabolitos tienen una actividad inmunomoduladora y por lo tanto pueden ser usados en el biocontrol de insectos (Jegorov et al., 1994). Otro metabolito es el ácido 2,6 piridin dicarboxílico, conocido comúnmente como ácido dipicolinico, y que mostró ser tóxico para ninfas de Bemisia argentifolii (Asaff et al., 2005).
Existen pocos estudios sobre el efecto que tienen los depsipéptidos sobre los enemigos naturales de los insectos plaga. Hu et al. (2009) demostraron que era insignificante la toxicidad de las destruxinas sobre Serangium japonicum, enemigo natural de Bemisia tabaci, sin embargo, se debe seguir avanzando en esta área de estudio para demostrar el efecto nulo de estas moléculas en insectos benéficos.
Respecto a los hongos que ocasionan enormes pérdidas económicas en la agricultura, destruyendo cultivos en el campo y en su almacenamiento, su control puede ser llevado a cabo a través de antibióticos y otros metabolitos con acción fungicida. Estos metabolitos actúan principalmente sobre la síntesis de la pared celular de los hongos, Los principales metabolitos que inhiben la síntesis del glucano son las equinocandinas (Figura 2) que dañan la pared celular de Aspergillus spp. en una acción fungistática (Denning, 2003), estas moléculas son hexapéptidos cíclicos N-acetilados con una cadena alifática de diferentes longitudes (Turner y Current, 1997), que son sintetizados por diversos hongos, cuya acción biológica se ha mejorado con modificaciones químicas (Vicente et al., 2003). En el caso de la biosíntesis del polímero de quitina, la enzima quitina sintasa es inhibida competitivamente por polioxinas y nicomicinas (Gabib, 1991), estos metabolitos han sido usados exclusivamente en la agricultura como fungicidas debido a su baja actividad biológica sobre hongos patógenos para el ser humano (Cohen, 1993).
En cuanto a los depsipéptidos cíclicos, se ha comprobado su acción antifúngica al probar el efecto de la destruxina B de M. anisopliae sobre el hongo fitopatógeno Alternaria brassicae, encontrándose que es susceptible a este metabolito (Pedras et al., 2002).
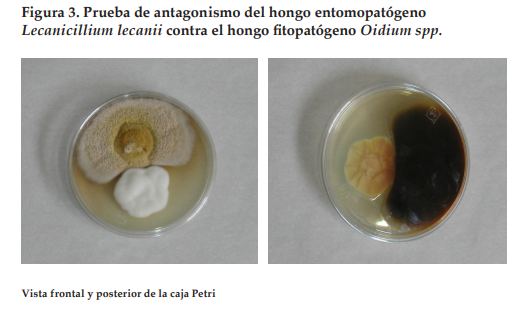
Los HE aunque son agentes de control de insectos plaga, suelen producir metabolitos que pueden tener acción fungicida al encontrarse en nichos con alta biodiversidad y competencia microbiana. En estudios preliminares (Figura 3) se ha mostrado su capacidad para inhibir el crecimiento miceliar de otros hongos; en la prueba de antagonismo entre el hongo entomopatógeno Lecanicillium lecanii y el hongo Oidium sp, conocido comúnmente como cenicilla del durazno, se observó su acción inhibitoria, sugiriendo que L. lecani produce enzimas o agentes antifúngicos que posiblemente afectan o inhiben la biosíntesis de la pared celular del hongo (Barranco et al., 2009).
Las enzimas y metabolitos mencionados pueden ser producidos por procesos fermentativos en cultivo líquido o sólido y ser escalados hacia una producción industrial. Los HE que generalmente se utilizan para el control de artrópodos pueden ampliar su uso contra hongos fitopatógenos abriendo un área de investigación en el campo de la agricultura (Anke y Antelo, 2009). Por lo que la búsqueda de nuevos compuestos bioactivos, seguida de la síntesis y optimización de análogos, desarrollaría un área de conocimiento que apoyaría el manejo integrado de insectos y hongos desde una perspectiva sustentable con productos como enzimas y toxinas; con lo que se crearía una nueva generación de bioinsecticidas de gran importancia biotecnológica y agrícola, que requerirá de un grupo de investigación multidisciplinario en México.
CONCLUSIONES
El control de insectos por los HE se lleva a cabo utilizando las esporas como la forma infectiva. En el proceso patogénico producen metabolitos que afectan y matan a los insectos como las enzimas, depsipéptidos cíclicos y toxinas. Sin embargo, estos metabolitos, junto con los antibióticos y otras moléculas que inhiben la síntesis de la pared celular de los hongos, también pueden emplearse para controlar las enfermedades en las plantas producidas por los hongos fitopatógenos. Esa dualidad de los HE sugiere la posibilidad de producir moléculas bioactivas que pueden desarrollarse en productos biotecnológicos con acción insecticidas o fungicidas, con lo cual se abriría un área agrobiotecnológica de investigación y desarrollo.
